

Dna Is Held Together By
Biologists in the 1940s had difficulty in accepting DNA as the genetic textile because of the apparent simplicity of its chemistry. Deoxyribonucleic acid was known to be a long polymer composed of only four types of subunits, which resemble i another chemically. Early on in the 1950s, Deoxyribonucleic acid was showtime examined by 10-ray diffraction assay, a technique for determining the three-dimensional diminutive structure of a molecule (discussed in Affiliate 8). The early on x-ray diffraction results indicated that DNA was composed of two strands of the polymer wound into a helix. The observation that DNA was double-stranded was of crucial significance and provided ane of the major clues that led to the Watson-Crick structure of Deoxyribonucleic acid. Only when this model was proposed did Dna's potential for replication and information encoding become credible. In this section nosotros examine the structure of the DNA molecule and explain in full general terms how it is able to shop hereditary data.
A DNA Molecule Consists of 2 Complementary Chains of Nucleotides
A DNA molecule consists of 2 long polynucleotide bondage composed of four types of nucleotide subunits. Each of these chains is known as a Deoxyribonucleic acid chain, or a Dna strand. Hydrogen bonds between the base of operations portions of the nucleotides hold the two chains together (Figure four-3). As nosotros saw in Chapter ii (Panel two-6, pp. 120-121), nucleotides are composed of a 5-carbon sugar to which are fastened one or more than phosphate groups and a nitrogen-containing base. In the case of the nucleotides in DNA, the carbohydrate is deoxyribose attached to a single phosphate grouping (hence the proper name deoxyribonucleic acrid), and the base may exist either adenine (A), cytosine (C), guanine (Grand), or thymine (T). The nucleotides are covalently linked together in a concatenation through the sugars and phosphates, which thus form a "courage" of alternating sugar-phosphate-sugar-phosphate (come across Figure 4-three). Because merely the base differs in each of the four types of subunits, each polynucleotide concatenation in DNA is analogous to a necklace (the backbone) strung with four types of chaplet (the iv bases A, C, G, and T). These same symbols (A, C, M, and T) are also commonly used to denote the 4 different nucleotides—that is, the bases with their fastened carbohydrate and phosphate groups.
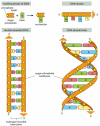
Figure 4-3
Dna and its building blocks. Dna is fabricated of four types of nucleotides, which are linked covalently into a polynucleotide concatenation (a DNA strand) with a sugar-phosphate backbone from which the bases (A, C, G, and T) extend. A DNA molecule is composed of ii (more...)
The fashion in which the nucleotide subunits are lined together gives a Dna strand a chemical polarity. If we call back of each sugar every bit a block with a protruding knob (the 5′ phosphate) on one side and a hole (the 3′ hydroxyl) on the other (see Figure 4-iii), each completed concatenation, formed by interlocking knobs with holes, volition have all of its subunits lined up in the same orientation. Moreover, the two ends of the chain will exist easily distinguishable, as one has a hole (the 3′ hydroxyl) and the other a knob (the 5′ phosphate) at its terminus. This polarity in a DNA chain is indicated past referring to one terminate as the 3′ end and the other equally the v′ stop.
The three-dimensional structure of DNA—the double helix—arises from the chemical and structural features of its 2 polynucleotide chains. Considering these two bondage are held together by hydrogen bonding between the bases on the different strands, all the bases are on the inside of the double helix, and the carbohydrate-phosphate backbones are on the outside (see Figure 4-three). In each example, a bulkier two-ring base (a purine; see Console ii-6, pp. 120–121) is paired with a single-band base of operations (a pyrimidine); A e'er pairs with T, and G with C (Effigy iv-4). This complementary base of operations-pairing enables the base of operations pairs to be packed in the energetically most favorable arrangement in the interior of the double helix. In this arrangement, each base pair is of similar width, thus holding the saccharide-phosphate backbones an equal distance autonomously along the DNA molecule. To maximize the efficiency of base-pair packing, the two saccharide-phosphate backbones wind effectually each other to class a double helix, with one complete turn every ten base pairs (Figure 4-5).

Figure 4-4
Complementary base pairs in the DNA double helix. The shapes and chemical structure of the bases allow hydrogen bonds to class efficiently only between A and T and between K and C, where atoms that are able to form hydrogen bonds (encounter Panel 2-iii, pp. 114–115) (more...)

Figure 4-five
The Dna double helix. (A) A space-filling model of 1.5 turns of the DNA double helix. Each turn of Dna is made up of x.four nucleotide pairs and the heart-to-center altitude between adjacent nucleotide pairs is 3.4 nm. The coiling of the ii strands around (more...)
The members of each base of operations pair can fit together within the double helix just if the ii strands of the helix are antiparallel—that is, only if the polarity of one strand is oriented opposite to that of the other strand (run across Figures 4-3 and 4-4). A event of these base-pairing requirements is that each strand of a DNA molecule contains a sequence of nucleotides that is exactly complementary to the nucleotide sequence of its partner strand.
The Structure of Dna Provides a Mechanism for Heredity
Genes bear biological information that must be copied accurately for transmission to the next generation each time a cell divides to form two daughter cells. Two cardinal biological questions arise from these requirements: how can the information for specifying an organism be carried in chemical form, and how is it accurately copied? The discovery of the structure of the Dna double helix was a landmark in twentieth-century biology because information technology immediately suggested answers to both questions, thereby resolving at the molecular level the problem of heredity. We discuss briefly the answers to these questions in this section, and we shall examine them in more detail in subsequent chapters.
Deoxyribonucleic acid encodes information through the guild, or sequence, of the nucleotides along each strand. Each base—A, C, T, or G—can be considered equally a alphabetic character in a 4-letter alphabet that spells out biological messages in the chemical structure of the Deoxyribonucleic acid. As nosotros saw in Affiliate ane, organisms differ from 1 some other because their respective DNA molecules have dissimilar nucleotide sequences and, consequently, carry dissimilar biological messages. But how is the nucleotide alphabet used to make messages, and what do they spell out?
As discussed above, information technology was known well before the structure of DNA was determined that genes comprise the instructions for producing proteins. The Deoxyribonucleic acid letters must therefore somehow encode proteins (Figure iv-vi). This relationship immediately makes the problem easier to empathize, because of the chemic character of proteins. Every bit discussed in Chapter three, the properties of a poly peptide, which are responsible for its biological function, are determined past its three-dimensional structure, and its structure is determined in turn by the linear sequence of the amino acids of which it is composed. The linear sequence of nucleotides in a gene must therefore somehow spell out the linear sequence of amino acids in a poly peptide. The exact correspondence betwixt the 4-letter nucleotide alphabet of DNA and the xx-letter amino acid alphabet of proteins—the genetic code—is not obvious from the Deoxyribonucleic acid construction, and it took over a decade later on the discovery of the double helix earlier it was worked out. In Chapter half dozen we draw this code in item in the form of elaborating the process, known as gene expression, through which a cell translates the nucleotide sequence of a factor into the amino acid sequence of a poly peptide.

Figure 4-vi
The human relationship between genetic information carried in DNA and proteins.
The complete set of information in an organism's Dna is called its genome, and information technology carries the information for all the proteins the organism will ever synthesize. (The term genome is also used to describe the Dna that carries this information.) The amount of data contained in genomes is staggering: for instance, a typical homo cell contains 2 meters of Dna. Written out in the four-letter of the alphabet nucleotide alphabet, the nucleotide sequence of a very minor human gene occupies a quarter of a page of text (Figure 4-vii), while the consummate sequence of nucleotides in the human genome would make full more than a thousand books the size of this one. In improver to other critical data, it carries the instructions for about 30,000 distinct proteins.

Figure 4-seven
The nucleotide sequence of the human being β-globin gene. This factor carries the information for the amino acid sequence of 1 of the 2 types of subunits of the hemoglobin molecule, which carries oxygen in the blood. A different factor, the α-globin (more...)
At each prison cell division, the cell must copy its genome to pass it to both daughter cells. The discovery of the structure of DNA as well revealed the principle that makes this copying possible: because each strand of DNA contains a sequence of nucleotides that is exactly complementary to the nucleotide sequence of its partner strand, each strand tin can act as a template, or mold, for the synthesis of a new complementary strand. In other words, if we designate the 2 DNA strands as S and Southward′, strand S can serve equally a template for making a new strand South′, while strand S′ can serve as a template for making a new strand Southward (Figure 4-8). Thus, the genetic information in DNA can be accurately copied past the beautifully simple process in which strand S separates from strand S′, and each separated strand then serves as a template for the production of a new complementary partner strand that is identical to its old partner.
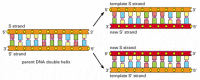
Effigy 4-8
Dna as a template for its own duplication. As the nucleotide A successfully pairs merely with T, and Chiliad with C, each strand of DNA can specify the sequence of nucleotides in its complementary strand. In this style, double-helical DNA can be copied precisely. (more...)
The ability of each strand of a DNA molecule to human activity as a template for producing a complementary strand enables a cell to copy, or replicate, its genes earlier passing them on to its descendants. In the side by side chapter we describe the elegant machinery the cell uses to perform this enormous chore.
In Eucaryotes, Deoxyribonucleic acid Is Enclosed in a Prison cell Nucleus
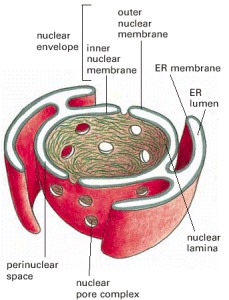
Nearly all the DNA in a eucaryotic cell is sequestered in a nucleus, which occupies near 10% of the total cell volume. This compartment is delimited past a nuclear envelope formed by 2 concentric lipid bilayer membranes that are punctured at intervals by big nuclear pores, which ship molecules between the nucleus and the cytosol. The nuclear envelope is directly connected to the extensive membranes of the endoplasmic reticulum. Information technology is mechanically supported by two networks of intermediate filaments: ane, called the nuclear lamina, forms a thin sheetlike meshwork within the nucleus, just below the inner nuclear membrane; the other surrounds the outer nuclear membrane and is less regularly organized (Effigy iv-9).
Figure 4-9
A cross-sectional view of a typical cell nucleus. The nuclear envelope consists of 2 membranes, the outer 1 being continuous with the endoplasmic reticulum membrane (run across also Figure 12-9). The infinite inside the endoplasmic reticulum (the ER lumen) (more...)
The nuclear envelope allows the many proteins that act on DNA to be concentrated where they are needed in the cell, and, every bit nosotros see in subsequent chapters, it too keeps nuclear and cytosolic enzymes split, a characteristic that is crucial for the proper functioning of eucaryotic cells. Compartmentalization, of which the nucleus is an example, is an of import principle of biology; it serves to establish an environment in which biochemical reactions are facilitated by the high concentration of both substrates and the enzymes that act on them.
Summary
Genetic data is carried in the linear sequence of nucleotides in DNA. Each molecule of Dna is a double helix formed from two complementary strands of nucleotides held together past hydrogen bonds between Thousand-C and A-T base pairs. Duplication of the genetic information occurs by the use of one DNA strand as a template for formation of a complementary strand. The genetic data stored in an organism's Dna contains the instructions for all the proteins the organism will ever synthesize. In eucaryotes, DNA is contained in the cell nucleus.

Dna Is Held Together By,
Source: https://www.ncbi.nlm.nih.gov/books/NBK26821/
Posted by: koneart1976.blogspot.com
0 Response to "Dna Is Held Together By"
Post a Comment